Lubna H Tahtamouni1 ![]() ,
Salem R Yasin1,
Reem N Abdellatif1,
Rema A Al-Khateeb1,
Zainab A Al-Mazaydeh1,
Samer I Al-Gharabli2,
Ali Z Elkarmi1
,
Salem R Yasin1,
Reem N Abdellatif1,
Rema A Al-Khateeb1,
Zainab A Al-Mazaydeh1,
Samer I Al-Gharabli2,
Ali Z Elkarmi1
For correspondence:- Lubna Tahtamouni Email: lubnatahtamuni@hu.edu.jo Tel:+96253903333
Received: 3 October 2016 Accepted: 24 December 2016 Published: 31 January 2017
Citation: Tahtamouni LH, Yasin SR, Abdellatif RN, Al-Khateeb RA, Al-Mazaydeh ZA, Al-Gharabli SI, et al. Inhibitory effect of Taraxacum officinale L (Compositae) aqueous root extract on spermatogenesis. Trop J Pharm Res 2017; 16(1):109-118 doi: 10.4314/tjpr.v16i1.14
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate if T. officinale root aqueous extract has anti-spermatogenic activity similar to that of the whole plant which was shown previously to inhibit spermatogenesis.
Methods: T. officinale aqueous extract was prepared by soaking 100 g of dried materials in 1 L distilled water for two days at 45 oC. Fifty adult male rats were divided into five groups and treated for 60 days. Four groups were gavaged with the whole plant or root aqueous extract in low or high doses. The male rat rats were allowed to mate with female rats. The control group received distilled water. Sperm count, motility and morphology as well as chromatin integrity were evaluated.
Results: Serum testosterone level, sperm parameters, pregnancy rate and average number of fetuses per pregnant females decreased significantly in the treated groups compared to control and in the root-receiving rats compared to the whole plant-receiving rats. Female rats which were mated with high dose root-receiving males did not deliver fetuses. Cross sections of seminiferous tubules of T. officinale-treated rats showed lesions and disorganized germinal epithelium. Late spermatogenesis maturation arrest (spermatid stage) was observed in all of the treated groups except the high dose root-receiving group which showed early maturation arrest (spermatocyte stage). In addition, the mRNA level of two spermatogonial stem cell markers responsible for self-renewal and proliferation of spermatogonia increased in high dose-receiving rats.
Conclusion: T. officinale root aqueous extract has inhibitory effects on spermatogenesis. Further studies are required to identify specific ingredient(s) in T. officinale that may be useful as male contraceptive(s).
Introduction
Treatment of disease began long ago with the use of herbs [1]. Taraxacum officinale (L.) Weber ex F.H. Wigg. (Compositae) is used in folk medicine as an anti-inflammatory, anti-oxidative, anti-carcinogenic and anti-coagulatory remedy [2]. Other health-promoting activities such as diuretic, laxative, cholagogue, anti-rheumatic, choleretic and hypoglycaemic have been attributed to the use of T. officinale [2].
In Jordan it is also used to enhance male fertility by enhancing sperm parameters such as concentration, motility and normal morphology. However, it has been proven recently that T. officinale whole plant or leaves aqueous extracts decrease male fertility instead of improving it by causing an imbalance between spermatogonia self-renewal and differentiation [3,4].
Spermatogonial stem cells (SSCs), which are located near the basement membrane of the seminiferous tubule [5], are responsible for the maintenance of spermatogenesis in males and are capable of producing mature sperms [6].
SSC self-renewal and differentiation are controlled by several markers [7]. Glial Cell-Derived Neurotrophic Factor (GDNF) is essential for self-renewal and proliferation of SSCs [8]. GDNF works through a multi-component receptor complex composed of GDNF family receptor alpha 1 (GFRα-1) and Ret (tyrosine kinase transmembrane protein) [9].
Promyelocytic Leukaemia Zinc-Finger (PLZF) expression is restricted to the undifferentiated spermatogonia and it prevents SSCs differentiation [10]. Macrophage Colony-Stimulating Factor (MCSF) is another stem cell marker secreted by the interstitial Leydig and myoid cells; its effect is restricted to undifferentiated spermatogonia [11].
The current work was conducted to investigate if T. officinale root aqueous extract has anti-spermatogenic activity similar to that of the whole plant. In addition, the work aimed at studying the effects of the root aqueous extract on GFRα-1, PLZF and MCSF spermatogonial stem cells (SSC) markers.
Methods
Plant collection and extract preparation
T. officinale was collected during the flowering season (February - October) of 2014 from Jubaiha region, Amman, Jordan. The plant was authenticated by a plant taxonomist at Raja’a Abu Eideh; The Hashemite University) and a voucher specimen (no. HU-42741) was deposited in the herbarium of the Department of Biology and Biotechnology, The Hashemite University, Zarqa, Jordan. The aqueous extract was prepared by soaking 100 g of air dried whole plant or root in 1 L of distilled water for two days at 45 °C. Afterwards, the mixture was filtrated by Whatman filter paper and then lyophilized [3,12].
Screening of chemical constituents of T. officinale
The determination of the chemical composition of T. officinale was performed by high performance liquid chromatography coupled with mass spectrometer (HPLC-MS). Briefly, 0.1 mg of each extract was dissolved in 1 ml of 20 % acetonitrile in water. Prepared samples were homogenized and centrifuged to avoid column blocking. Samples were introduced to HPLC-MS and photodiode array detector (Finigan Surveyor PDA Plus Detector, Thermo Scientific, USA). RP/C18 column (150 mm × 4.6 mm) at a flow rate of 1 ml/min was used. HPLC-MS spectra were collected for the extracts and compared to the data found in literature. As such, assignment of the chemical composition was possible only for those compounds which were already extracted and characterized for T. officinale.
Animal treatment
Fifty adult Wistar male rats (Rattus norvegicus) weighing between 150 and 200 g were randomly selected and housed individually in cages. The animals were maintained under standard conditions; tap water and feed were provided ad libitum. After one week of acclimatization [12], the rats were randomly divided into five different groups, each of ten animals. The animals of the first and second groups were gavaged with 1/20 and 1/10 of T. officinale whole plant aqueous extract LD50 and were considered the low dose whole plant receiving-group (LDWP) and the high dose whole plant-receiving-group (HDWP), respectively [3]. Rats of the third and fourth groups received 1/20 (3.68 g/kg) and 1/20 (1.84 g/kg) of the root aqueous extract LD50 and were considered the low dose root-receiving group (LDR) and the high dose root-receiving group (HDR), respectively. The fifth group of animals received distilled water and was considered the control group. The doses were given orally using a gavage needle for 60 consecutive days [12].
Fertility test
On the 55th day of treatment, each male rat was cohabitated with two adult pro-estrus female rats. The presence of a typical mating plug and/or sperms in the vaginal smear were considered indicators of successful mating. The number, weight and sex of the offspring were determined [12].
Male rat sacrifice
Male rats were sacrificed by cervical dislocation, and the body weight was recorded. Rats were dissected and liver, testes, kidneys and seminal vesicles were removed and weighed. Relative organ weight was calculated by dividing the absolute organ weight (g) by the body weight of rat on sacrifice day (g) multiplied by 100. Blood was collected by heart puncture in a plain blood tube and centrifuged at 2000 rpm for 10 min.
Sperm collection
Sperm samples were collected from cauda epididymis based on the method of Soleimanzadeh and Saberivand [13]. The samples were used to study sperm count, motility and morphology [14]. In addition, epididymal sperm samples were used to study chromatin integrity by acridine orange staining [15] and agarose gel electrophoresis. For acridine orange staining, two slides were prepared for each rat and at least two hundred sperms for each slide were counted [15].
Testosterone assay
Serum testosterone level was evaluated using the Immulite 1000 immunoassay system (Siemens, Germany). Briefly, serum from control or treated rats and alkaline phosphatase-conjugated testosterone were incubated in a test tube containing polystyrene beads coated with testosterone-specific antibodies. Serum testosterone competed with the enzyme-labeled testosterone for the limited number of antibody binding sites on the beads. A chemiluminescence substrate was added to the test tube where it was hydrolyzed by the alkaline phosphatase and the absorbance was measured spectrophotometrically (Siemens, Germany).
Histological studies
One of the testes was cut into small pieces (5 × 5 mm) and fixed in Bouin’s fixative. The samples were then dehydrated in ascending alcohol series and cleared in xylene. The tissues were imbedded in pure paraffin wax and serial sections of 6 µm were made and stained with hematoxylin and eosin. Finally, the sections were cleared and mounted in DPX and then were examined by Nikon Eclipse 50i microscope.
RNA extraction and quantitative real-time polymerase chain reaction (qRT-PCR)
For quantifying mRNA level of each SSC marker [11,16], quantitative real-time polymerase chain reaction (qRT-PCR) on Line Gene 9680 BioGR thermal cycler (Bioer Technology, USA) was carried out [17]. The SV Total RNA Isolation System (Promega, USA) was used for testicular RNA extraction and cDNA was prepared using Power cDNA synthesis kit (iNtRON Biotechnology). The quantification of the SSC markers (mcsf, gfra1, plzf) and b-actin (as a house keeping gene) was performed using the 2x Universal KAPA SYBR FAST qPCR Kit Master Mix (KAPA BIOSYSTEMS, USA). Each qRT-PCR was run as triplicate for each primer.
Ethical approval
The study was approved by the Institute Review Board (IRB) of The Hashemite University (AM/16/10/222/140115). Animal care, handling, and the animal experiments performed were approved by The Hashemite University Institutional Animal Care and Use Committee which conforms to the guidelines of National Research Council of the National Academics [18].
Statistical analysis
Statistical analysis of data was performed using STATISTICA 7 analysis program (StatSoft Inc., OK, USA). In order to determine differences between three or more means, one-way ANOVA with Fisher's LSD for multiple comparisons post-tests were performed. Data are presented as mean ± SD and the level of significance was set at p < 0.05.
Results
Phytochemical constituents of T. officinale
HPLC-MS was used to detect the chemical compounds present in T. officinale aqueous extracts. Various compounds were detected in the root aqueous extract including phenols (chlorogenic acid and chicoric acid), sesquiterpenes (tetrahydroridentin B, taraxa-colide-O-b glucopyranoside, ainslioside and 11b-13-dihydrolactucin), triterpene (taraxasterol and a and b amyrin) and coumarins (scopoletin and esculetin). The whole plant aqueous extract contained all of the above mentioned compounds in addition to hydroxycinnamic acid (caffeic acid) and monocaffeoyltartaric acid (phenols), luteolin 7-O-glucoside, quercetin 7-O-glucoside, luteolin 7-O-rutinoside and luteolin 7-diglucosides (flavonoid glycosides), ixerin D, taraxinic acid b-D-glucopyranoside and 11,13-dihydrotaraxinic acid (sesquiterpenes), and cichoriin and aesculin (coumarins).
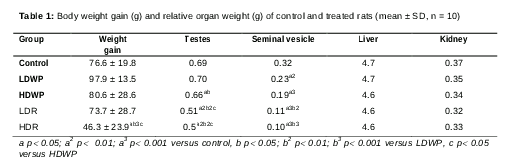
Body and organ weight of male rats
A significant decrease in body weight of the HDR-receiving rats was observed when compared to the control and whole plant-receiving rats (). In addition, there was a significant decrease in testis relative weight of the HDWP-; LDR- and HDR-receiving rats when compared to the control rats, and of the root-receiving rats when compared to the whole plant-receiving rats (). Similarly, the relative weight of the seminal vesicle showed a significant decrease in all of the treated groups when compared to control group, and in the root-receiving groups when compared to the LDWP-receiving group (). The relative weights of the liver and the kidney did not show a significant change in any of the treated groups as compared to the control ().
Serum testosterone level
T. officinale aqueous extract administration caused a significant decrease in serum testosterone level in all of the treated groups [LDWP: 1.6 ± 0.4 ng/mL (p < 0.05); HDWP: 1.4 ± 0.3 ng/mL (p < 0.05); LDR: 0.9 ± 0.1 ng/mL (p < 0.01); HDL: 0.3 ± 0.1 ng/mL (p < 0.001)] when compared to the control group (3.7 ± 1.2 ng/mL), in the root-receiving groups when compared to the whole plant-receiving groups (p < 0.05 for LDR and p < 0.01 for HDR), and in HDR-receiving group when compared to LDR-receiving group (p <0.05).
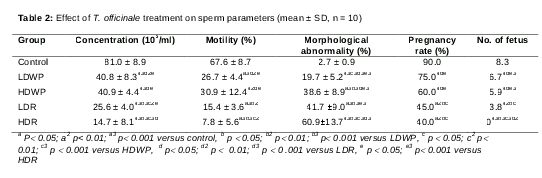
Sperm concentration, motility and morphology
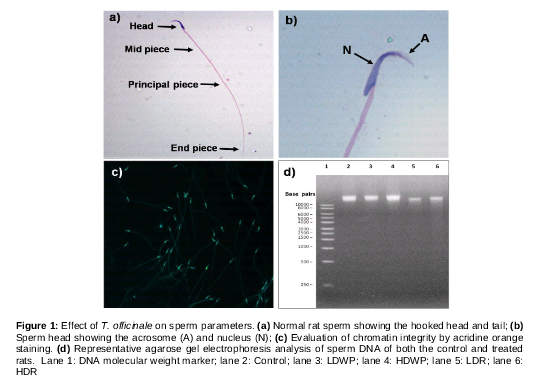
Sperm concentration and progressive motility in all of T. officinale-treated groups decreased significantly when compared to the control, and in the root-receiving groups as compared to the whole plant-receiving groups (). A rat sperm with normal morphology has an elongated tail composed of midpiece, principal piece and end piece, with intact hook-shaped head (a, b). The percentage of sperm morphological abnormalities increased significantly in the treated groups when compared to the control group, and in the root-receiving groups when compared to the whole plant-receiving groups ().
Sperm chromatin integrity
Sperm samples from cauda epididymis of the control and treated rats were stained with acridine orange. The results did not show DNA damage in either the control or treated samples as all sperms fluoresced green after excitation (c). A single intact band was observed in all of the lanes (control and treated groups), while the ladder pattern of DNA, which is an indication of fragmented DNA, was not observed (d).
Effect of T. officinale on pregnancy parameters
A significant decrease was observed in the pregnancy rate and the average number of fetuses/pregnant females in all of the treated groups as compared to the control group, and in the root-receiving groups when compared to the whole plant-receiving groups (). None of the female rats that were mated with the HDR-receiving males delivered fetuses after the regular pregnancy period (21 days) ().
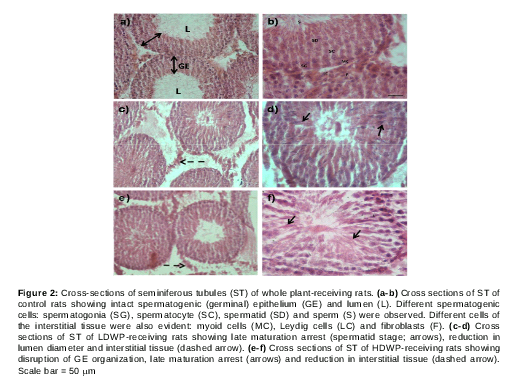
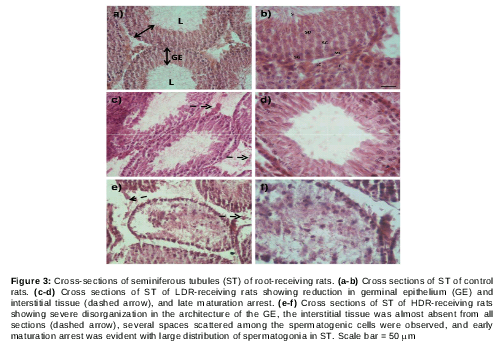
Effect of T. officinale on rat testicular histology
Microscopic examination of testicular cross sections of control rats showed normal histology (a, b). The germinal epithelium (GE) was intact and the lumen (L) was wide and full of sperms (a). In addition, different stages of sperm development were noticed (b). Cells of the interstitial tissue such as Leydig cells (LC), myoid cells (MC) and fibroblasts (F) were also observed (b).
However, testicular sections of LDWP-receiving group showed late maturation arrest (spermatid stage) (c, d; arrow) in most of the seminiferous tubules (ST) with a decreased number of sperms in the lumen and reduction in interstitial tissues (dashed arrow). HDWP-receiving group testicular sections showed reduction in ST numbers, most of which showed late maturation arrest, disorganized germinal epithelium and a reduction in the interstitial tissue (dashed arrow) (e, f). The LDR-receiving group ST cross sections showed late maturation arrest (arrows) with a reduction in germ cell number (reduced germinal epithelium thickness) and interstitial cells (dashed arrows) (c, d). On the other hand, cross sections prepared from seminiferous tubules of HDR-receiving rats showed sever disruption in ST architecture and large spaces between germ cells. In addition, early maturation arrest (spermatocyte stage) and germ cell hypoplasia were observed with almost a complete absence of interstitial tissue (dashed arrows). One striking observation in the HDR-receiving group was the absence of a clear lumen (e, f).
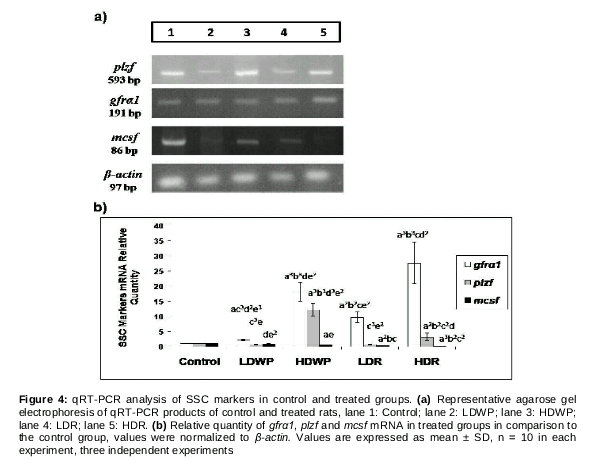
Gene expression of SSC markers
Three spermatogonial stem cell (SSC) self-renewal and proliferation genes were analyzed by qRT-PCR (a). The mRNA levels of these markers (gfra1, mcsf and plzf) were normalized to β-actin. The level of gfra1 mRNA increased significantly in all the treated groups when compared to the control, and in the HD-receiving groups when compared to the LD-receiving groups (b). plzf mRNA level increased significantly in the HD-receiving groups when compared to the control and the LD-receiving groups, and in the HDWP-receiving group when compared to the HDR-receiving group (b). On the other hand, mcsf mRNA level decreased significantly in all treated groups (except the LDWP-receiving group) when compared to the control group, and in the root-receiving groups when compared to the whole plant-receiving groups (b).
Discussion
Taraxacum officinale has been used in folk medicine for its different medicinal properties [2]. In addition to its known health benefits [1,2], T. officinale whole plant decoction has been traditionally used in Jordan to enhance sperm parameters such as concentration and motility [13]. However, recent studies have proved that the whole plant or leaves aqueous extracts decrease male fertility instead of improving it [3,4]. Thus, we aimed to investigate the effects T. officinale root aqueous extract on fertility parameters and spermatogonial stem cell (SSC) markers by comparing it to the whole plant aqueous extract.
T. officinale treatment caused a significant decrease in testis and seminal vesicle relative weights in treated groups (). Similar results were observed when male rats were treated with T. officinale whole plant or leaves aqueous extracts [3,4] and Echinops echinatus (Compositae) roots [19]. The decrease observed was attributed to a reduction in testosterone level.
Testosterone regulates spermatogenesis, sperm differentiation and induces the development of male accessory reproductive organs [20]. Thus, any irregularities in testosterone production and concentration would have detrimental effects on spermatogenesis. T. officinale aqueous extract caused a significant decrease in serum testosterone level in all the treated groups when compared to the control group and in the root-receiving rats when compared to the whole plant-receiving rats. This is correlated with the decrease in the interstitial tissue (the source of testosterone) observed in testicular sections of whole plant- or root-treated groups.
In our study, cauda epididymal sperm concentration, motility and normal morphology showed a significant decrease in all of T. officinale-treated groups when compared to the control group (). This result is in agreement with the results of Padashetty and Mishra [19]; their explanation was a reduction in serum testosterone level which is similar to our results.
The testicular histology of treated rats could provide another explanation for the reduction in sperm concentration observed. Testicular cross sections showed lesions in the seminiferous tubules, disorganized germinal epithelium, spermatogenesis maturation arrest and reduction in the interstitial tissue ( and ). The reduction in testosterone level disrupts spermatogenesis and reduces fertility parameters [21]. These results were similar to the results obtained by others [22].
Sperms with rapid swimming abilities and normal morphology have a fertilizing advantage over slower abnormal sperms [23]. T. officinale treatment reduced sperm parameters causing a reduction in the fertility rate and the number of fetuses/pregnant female which is in agreement with previously published results of Calendula officinalis (Compositae) administration [24].
The current study measured the mRNA levels of three spermatogonial stem cell (SSC) markers: GDNF family receptor alpha 1 (gfrα1), macrophage colony-stimulating factor (mcsf) and promyelocytic leukaemia zinc-finger (plzf) by qRT-PCR (). These markers induce self-renewal and proliferation of undifferentiated spermatogonia rather than differentiation [7]. GDNF and its receptor GFRα1 are necessary for the maintenance and proliferation of undifferentiated spermatogonia. Over-expression of these markers led to accumulation of spermatogonia that are unable to differentiate in addition to testicular atrophy [25]. Hofmann [26] suggested that mice over-expressing gfra1 are infertile, develop testicular tumors and accumulate spermatogonia.
PLZF exerts its effects on undifferentiated spermatogonia only and it interacts with GDNF to block spermatogonia differentiation [27]. Costoya et al. [28] showed that testes lacking PLZF have an increased level of spermatogonia apoptosis. A nonsense mutation in plzf gene showed defective spermatogenic self-renewal [29]. Silvan et al [30] reported that MCSF interacts with GDNF and acts specifically on undifferentiated SSCs and that its effect causes spermatogonia self-renewal rather than differentiation.
The reduced expression of mcsf () could be explained by the histology of the seminiferous tubules of the T. officinale-treated rats -particularly the high dose root-receiving animals, as there was a lack of interstitial tissue (the source of MCSF) from the treated animals (). On the other hand, the over-expression of gfra1 and plzf () might have led to enhanced self-renewal of spermatogonia as was noticed in the testicular sections of HD-treated groups (). This enhancement of self-renewal was at the expense of differentiation induction and sperm production. In turn, this might have contributed to the disruption in spermatogenesis and the maturation arrest observed.
Taraxacum officinale is rich in phytochemicals. Nevertheless, few studies investigated the effects of individual compounds identified in T. officinale extracts on spermatogenesis. Previous studies have shown that flavonoids and amyrin inhibited spermatogenesis [31-33]. On the other hand, chlorogenic acid was found to stimulate spermatogenesis by inhibiting testosterone degradation [34]. However, the difference in the effects of the two extracts (root and whole plant) cannot be explained based on these compounds since these compounds were found in both the root and whole plant aqueous extracts. Studying the effects of individual phytochemical constituent of T. officinale aqueous extract on spermatogenesis is beyond the scope of this study.
Conclusion
T. officinale root and whole plant aqueous extracts have been shown to exert inhibitory effects on spermatogenesis. More studies should be conducted to identify the specific anti-spermatogenic active ingredient(s) and whether the anti-fertility actions are reversible or not. Furthermore, additional studies are advised to investigate the specific effect of T. officinale aqueous extracts on spermatogonial stem cell markers’ regulatory pathways. These studies could lead to the identification of specific ingredient(s) in T. officinale that might be used as male contraceptive.
Declarations
Acknowledgement
References
Archives


News Updates